[Tham khảo:
+”Hóa Sinh Y Học”, ĐHYD TPHCM.
+”Essential Cell Biology”, chapter 16, 4th edition
+”Hormone-Receptor: http://www.yhoccanban.com/2013/03/hormone-receptor.html].
……………………………………………………………………………………………………………………………………………………..
I. Vị trí tiếp nhận của hormone và receptor:
1/ Receptor trên màng tế bào: tiếp nhận hormone có bản chất peptide, protein, catecholamine (như glucagon, insulin, epinephrine,…)
2/ Receptor trong bào tương: tiếp nhận hormone có bản chất steroid (như testosterone, estrogen, retinoid acid,…)
3/ Receptor trong nhân: tiếp nhận hormone T3, T4 của tuyến giáp.
Ở đây ta chủ yếu đề cập các cơ chế truyền tín hiệu cần chất truyền tin thứ hai (second messenger) nên chỉ xét các receptor trên màng tế bào.
II. 3 loại thụ thể chính trên màng tế bào:
- Receptor liên kết kênh ion:
+Chuyển tín hiệu hóa học thành tín hiệu đuện.
+Thường gặp ở cơ chế truyền tín hiệu thần kinh.
- Receptor liên kết protein G (receptor 7-helix):
+Họ receptor lớn nhất trong các receptor trên màng.
+Protein G truyền tín hiệu từ receptor đến protein đáp ứng. Protein G có cấu trúc trimer với 3 bán đơn vị alpha, beta, gamma. Bán đơn vị alpha có khả năng liên kết với GDP (khi đó bất hoạt) hoặc GTP (khi đó được hoạt hóa), có hoạt tính GTPase.

Source: http://www.web-books.com/MoBio/Free/Ch6D2.htm

Source: https://signaltransductionpathway.weebly.com/g-protein.html
+Cơ chế: Chất dẫn truyền bám vào mặt ngoài receptor làm biến đổi cấu trúc receptor, tạo điều kiện thuận lợi cho mặt trong tiếp nhận protein G. Khi đó GDP ở tiểu phần alpha được biến đổi thành GTP, trước khi có sự phân ly cấu trúc trimer thành tiểu phần alpha và đơn vị beta-gamma. Hai thành phần này sau đó thể hiện cơ chế tác động của nó lên các chất khác.

Source: https://courses.washington.edu/conj/bess/gpcr/gpcr.htm
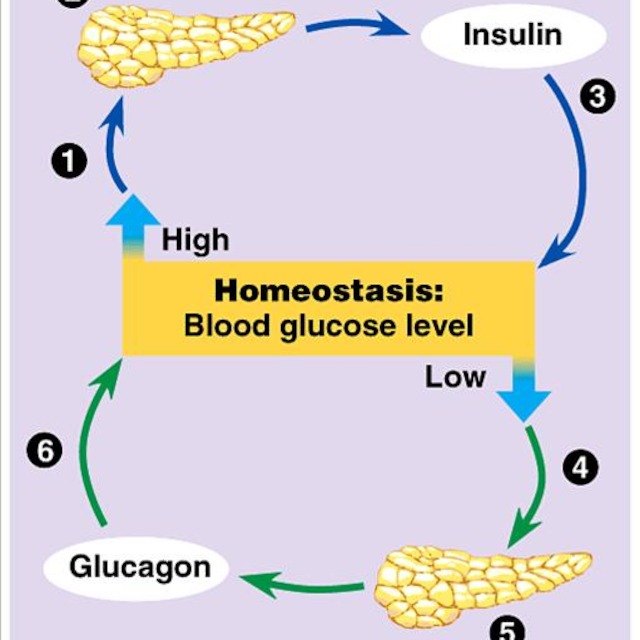
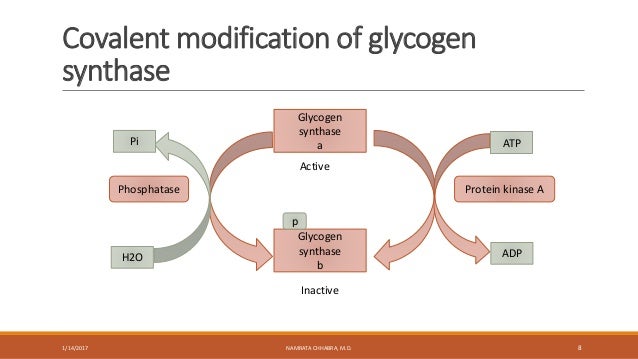
Ví dụ: tiểu phần alpha có thể gắn vào enzyme adenyl cyclase, kích thích tổng hợp chất truyền tin thứ hai là cAMP, hoạt hóa protein kinase A tham gia các quá trình chuyển hóa. Chẳng hạn protein kinase A có thể tiếp tục hoạt hóa phosphorylase kinase, từ đó phosphoryl hóa glycogen phosphorylase, tăng phân giải glycogen.
Tiểu phần alpha thường tác động lên enzyme thì đơn vị beta-gamma lại thường nhắm đến kênh ion:

Source: https://courses.washington.edu/conj/bess/gpcr/gpcr.htm
Đơn vị beta-gamma được hoạt hóa có khả năng mở kênh K+ trên màng bào tương, tăng tính thấm của màng đối với K+, từ đó làm giảm nhịp tim ở tế bào cơ tim.
+Phân loại protein G: gồm có Gs chiếm đa số (hoạt hóa enzyme adenyl cyclase hoặc tác động kênh ion), Gi (ức chế adenyl cyclase) và Gq (hoạt hóa enzyme phospholipase C).
- Receptor liên kết enzyme (receptor 1-helix):
+Protein xuyên màng.
+Ở mặt trong có những miền hoạt động như enzyme dị lập thể, thường gặp nhất là enzyme Tyrosine.
+Khi hormone gắn với receptor (ví dụ như insulin ở P1), enzyme kinase nội bào tự phosphoryl hóa nhờ ATP, từ đó phosphoryl hóa Tyrosine của protein khác, từ đó truyền tín hiệu đi như dòng thác tín hiệu.

Source: http://membranereceptors.com/transduction-process/enzyme-linked-receptors/
III. Các con đường truyền tin chủ yếu:
Đó là các lộ trình qua Tyrosine kinase (liên kết thác tín hiệu Ras), adenyl cyclase (chất truyền tin thứ hai là cAMP) hay phosphoinositid (chất truyền tin thứ hai là Inositol 1,4,5-triphosphate được tạo từ sự thủy phân phosphoinositid nhờ phospholipase C xúc tác). Tóm tắt cơ chế ở lần lượt các hình sau:
*Lộ trình Tyrosine kinase:

Source: https://www.researchgate.net/figure/The-function-of-Ras-and-its-deregulation-Ras-is-an-essential-component-of-the-signaling_fig4_26685709
Ras là loại protein có thể mang GDP hay GTP, tương ứng với trạng thái “im lặng” hay “hoạt động”. Khi một loại protein (adaptor protein) được phosphoryl hóa nhờ receptor Tyrosine kinase thì nó dẫn tới sự hình thành nhân tố chuyển GDP của Ras thành GTP (nhân tố này là Ras guanine nucleotide exchange factor, hay Ras-GEF) từ đó chuỗi tín hiệu truyền đi, vào trong nhân kích thích tăng sinh, biệt hóa hay sinh tồn. Hình trên cho thấy những tổn hại trong lộ trình trên có thể gây ung thư (cancer).
Tổng quát, receptor liên kết enzyme thường tiếp nhận các chất tín hiệu liên quan quá trình sinh trưởng, và dĩ nhiên sự đột biến trong truyền tin gắn liền ung thư.
*Lộ trình adenyl cyclase:
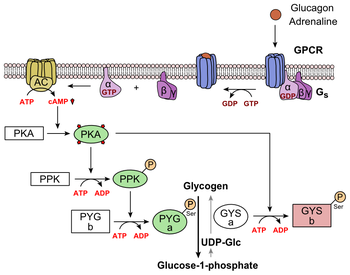
Source: https://en.wikipedia.org/wiki/Glucagon
(Cơ chế này ở P1 cũng có nói rồi, bản chất là protein G, cụ thể hơn là tiểu phần alpha, tác động lên adenyl cyclase, cuối cùng dẫn đến kích thích phân giải glycogen).
*Lộ trình phosphoinositid:

Source: http://courses.washington.edu/conj/gprotein/ip3.htm
Protein G, cụ thể hơn là tiểu phần alpha loại protein Gq có tác động đến enzyme phospholipase C. Enzyme này có thể thủy phân phosphoinositid thành Inositol 1,4,5-triphosphate (hay còn gọi là IP3) và đây là chất truyền tin thứ 2, có thể gắn vào các kênh Ca2+ phụ thuộc phối tử (Ligand-gated Ca2+ channel) trên màng lưới nội cơ tương, giải phóng Ca2+ ra tế bào chất, nơi mà nó tham gia nhiều quá trình chuyển hóa khác.